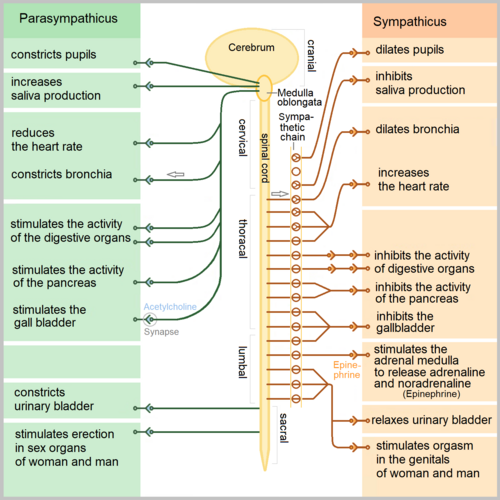
The sympathetic division coordinates the organism's response to stress, exercise, and emergency. Its anatomical architecture reflects this function: preganglionic neurons originate in a restricted spinal cord segment, diverge widely to reach paravertebral and prevertebral ganglia close to the spinal column, and from there project long postganglionic axons to widely distributed target organs. This organization allows a small number of preganglionic neurons to drive a large-scale, coordinated sympathetic response that simultaneously increases cardiac output, redistributes blood flow to skeletal muscle, dilates bronchi, and mobilizes fuel stores.
Thoracolumbar Preganglionic Origin. Sympathetic preganglionic neurons have their cell bodies in the intermediolateral cell column (also called the lateral horn) of the spinal cord gray matter, spanning from the first thoracic (T1) to the second or third lumbar (L2-L3) spinal segments. This is why the sympathetic division is termed "thoracolumbar." The myelinated preganglionic axons exit the spinal cord via the ventral roots, enter the spinal nerves briefly, and then leave via white rami communicantes to reach the sympathetic chain ganglia. The preganglionic fibers are relatively short compared to their parasympathetic counterparts, because the sympathetic ganglia are located close to the spinal column rather than at or near the target organ.24
Paravertebral (Sympathetic Chain) Ganglia. The paravertebral ganglia form two paired chains running bilaterally along the lateral surfaces of the vertebral column from the base of the skull to the coccyx. They are connected to each other by interganglionic rami, creating the sympathetic trunk or paravertebral chain. There are typically 22 to 23 ganglia per chain: 3 cervical, 10 to 12 thoracic, 4 lumbar, and 4 to 5 sacral, plus the ganglion impar at the coccyx where the two chains fuse. A preganglionic fiber entering the sympathetic chain via a white ramus communicans can take one of three routes: it may synapse with a postganglionic neuron at the same level, ascend or descend within the chain to synapse at a different level (which is how preganglionic fibers from thoracic levels T1 through T4 [T1-T4] reach the superior, middle, and inferior cervical ganglia to innervate the head, neck, and heart), or pass through the chain without synapsing to reach a prevertebral ganglion via a splanchnic nerve. Postganglionic fibers leaving the chain return to the spinal nerve via gray rami communicantes and travel with the somatic nerves to their peripheral targets.24
Prevertebral (Collateral) Ganglia. The prevertebral ganglia lie anterior to the vertebral column in the abdominal cavity, clustered around the origins of major abdominal arteries. The three principal prevertebral ganglia are the celiac ganglion (receiving input primarily from thoracic levels 5 through 12 [T5-T12] via the greater splanchnic nerve), the superior mesenteric ganglion (thoracic 10 through lumbar 1 [T10-L1] via the lesser splanchnic nerve), and the inferior mesenteric ganglion (lumbar 1 through 3 [L1-L3] via the lumbar splanchnic nerves). Preganglionic fibers reach these ganglia via the splanchnic nerves, which pass through the diaphragm without synapsing in the paravertebral chain. Postganglionic fibers from prevertebral ganglia innervate the abdominal and pelvic viscera, including the stomach, small intestine, large intestine, liver, pancreas, kidneys, adrenal glands, bladder, and reproductive organs. The long preganglionic fibers of the splanchnic nerves are surgically accessible, and splanchnic nerve block is used clinically for intractable visceral pain from pancreatic cancer and other abdominal malignancies.2
The Adrenal Medulla as a Modified Sympathetic Ganglion. The adrenal medulla is embryologically derived from neural crest cells that migrated to the adrenal gland and differentiated into chromaffin cells rather than classic postganglionic neurons. It is functionally homologous to a sympathetic ganglion: preganglionic sympathetic fibers travel via the greater splanchnic nerve to synapse directly on chromaffin cells using acetylcholine (ACh) at nicotinic ganglionic (NN) receptors, bypassing the usual postganglionic neuron entirely. Rather than releasing norepinephrine (NE) at a neuromuscular junction, chromaffin cells secrete a mixture of epinephrine (approximately 80%) and norepinephrine (approximately 20%) directly into the bloodstream, functioning as an endocrine organ. Epinephrine release from the adrenal medulla produces widespread sympathetic effects, with preferential activation of beta-2 adrenergic receptors (beta-2 ARs) in bronchial smooth muscle and peripheral vasculature because circulating epinephrine reaches these receptors in concentrations sufficient to engage beta-2 ARs, which have higher epinephrine affinity than the neurally released norepinephrine that dominates at alpha-1 adrenergic receptor (alpha-1 AR)-rich vascular beds. This distinction between neurally released NE (acting locally at the neuroeffector junction) and circulating epinephrine (acting via the bloodstream) is pharmacologically fundamental and explains why the adrenal medulla response has a different hemodynamic signature from direct sympathetic nerve activation.25
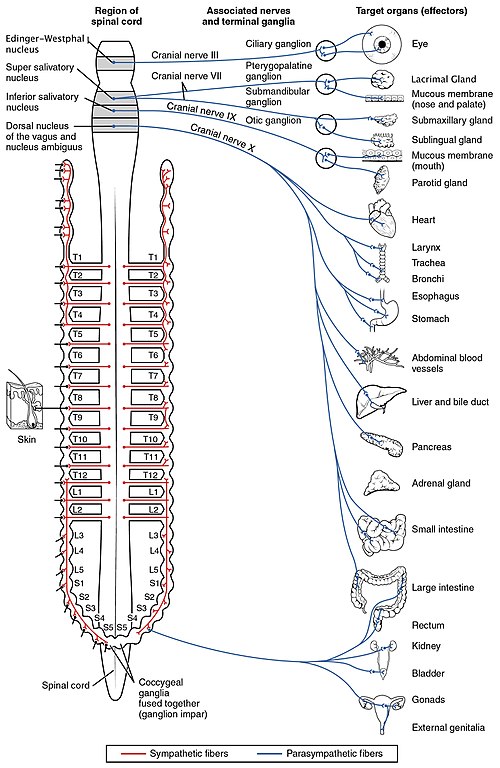
The cervical sympathetic chain (superior, middle, inferior/stellate ganglia) is clinically accessible: stellate ganglion block is used for complex regional pain syndrome, refractory ventricular arrhythmias, and hyperhidrosis. Horner syndrome (ptosis, miosis, anhidrosis, enophthalmos) results from interruption of the oculosympathetic pathway at any level from hypothalamus to superior cervical ganglion to iris dilator; localizing the lesion requires understanding the three-neuron oculosympathetic arc. The long preganglionic fibers of the splanchnic nerves make splanchnic nerve block feasible for visceral pain control. The adrenal medulla, innervated directly by preganglionic fibers, is the target in pheochromocytoma: catecholamine excess from chromaffin cell tumors produces hypertensive crises that must be managed with alpha-blockade before beta-blockade to avoid unopposed alpha-mediated vasoconstriction.